Как появились митохондрии согласно теории симбиогенеза. Происхождение эукариот
Следует отметить три гипотезы происхождения эукариотических клеток:
- симбиотическая гипотеза , или симбиогенез ,
- инвагинационная ,
- химерная .
На сегодняшний день в научном мире основной гипотезой происхождения эукариот признается симбиогенез .
Согласно симбиогенезу такие органеллы эукариотических клеток как митохондрии , хлоропласты и жгутики произошли путем внедрения одних прокариот в другую, более крупную прокариотическую клетку, сыгравшую роль клетки-хозяина.
В симбиотической гипотезе есть трудности при объяснении происхождения ядра эукариотических клеток и в вопросе, какой же все-таки прокариот выступил хозяином. Данные молекулярного анализа генома и белков эукариот показывают, что, с одной стороны, это был организм близкий к археям (раньше относились к бактериям, потом их выделили в отдельную ветвь). С другой стороны, в эукариотах имеются белки (и ответственные за их синтез гены), характерные для совершенной других групп прокариот.
Согласно инвагинационной гипотезе происхождения эукариотических клеток их органоиды образовались путем впячивания цитоплазматической мембраны с последующим отделением этих структур. Образовывались что-то вроде шариков, окруженных мембраной и содержащих внутри цитоплазму и захваченные сюда соединения и структуры. В зависимости от того, что попало внутрь, сформировались разные органоиды.
У прокариот нет настоящих органелл, их функции как раз и выполняют впячивания мембраны. Поэтому легко представить подобное ее отшнуровывание. Также в пользу инвагинационной гипотезы говорит схожесть цитоплазматической мембраны и двойных мембран органелл.
С точки зрения инвагинагенеза происхождение ядра легко объяснимо, но необъяснимо, почему геном и рибосомы ядерно-цитоплазматического комплекса отличаются от таковых в хлоропластах и митохондриях (вспомним, что в них также есть ДНК и рибосомы). Причем в указанных органеллах система биосинтеза белка (ДНК, РНК, рибосомы) схожа с прокариотами.
Это отличие хорошо объяснимо как раз с точки зрения первой, симбиотической, гипотезы. Согласно ей в анаэробный прокариот так или иначе попал аэробный прокариот. Он не переварился, а стал, наоборот, питаться за счет клетки-хозяина. В свою очередь он использовал кислород для получения энергии, а этот способ окисления намного эффективнее, избыток энергии он отдавал хозяину-прокариоту, который в таком случае также получал выгоду. Возник симбиоз. В последствии внедрившийся прокариот упростился, часть его генома мигрировала в клетку-хозяина, он уже не мог существовать независимо.
Подобным образом симбиогенез объясняет происхождение хлоропластов. Только внедрялись уже прокариоты, способные к фотосинтезу (подобные синезеленым водорослям).
Первые эукариоты без фотосинтезирующих симбионтов дали начало животным, у которых они появились - растениям.
В настоящее время существуют простейшие (одноклеточные эукариотические организмы) у которых нет митохондрий или хлоропластов. Зато вместо них в цитоплазме поселяются прокариоты-симбионты, выполняющие соответствующие функции. Этот факт, а также схожесть системы биосинтеза белка митохондрий и пластид с прокариотами рассматриваются как доказательства симбиогенеза. Доказательством также служит то, что митохондрии и хлоропласты размножаются самостоятельно, они никогда не строятся клеткой с нуля.
В пользу третьей, химерной, гипотезы происхождения эукариотических клеток, говорит большой размер их генома, который превосходит бактериальный в тысячи и более раз, а также разнообразие синтезируемых белков, встречающихся в разных группах прокариот. Понятно, что на протяжении эволюции эукариот их геном усложнялся, он удвоился, в нем появилось множество регулирующих генов. Но все же первоначальное увеличение размера генома могло произойти за счет объединения геномов нескольких прокариот.
Возможно в древности некий прокариот приобрел способность к фагоцитозу и, питаясь таким образом, поглощал в том числе других прокариот, которые не всегда переваривались. Их геном содержал полезные для хозяина гены, и он включал их в свой геном. Возможно некоторые из оказавшихся внутри прокариот становились органеллами, что объединяет химерную гипотезу с симбиогенезом.
Поскольку все перечисленные гипотезы имеют сильные и слабые стороны, а также во многом не исключают положения друг друга, то, на наш взгляд, в происхождении структур эукариотических клеток могло сыграть роль сочетание множества факторов, описываемых разными гипотезами.
Следует также отметить, что согласно симбиотической гипотезе происхождение других мембранных органелл таких как вакуоли, комплекс Гольджи и др. можно рассматривать как дальнейшее упрощение, например, митохондрий.
Также отметим, что сочетание в эукариотах белков из разных прокариот вовсе может не указывать на химеризм первых. Возможно, что определенные ферментативные функции могут выполнять только белки конкретного строения. И эволюция эукариот повторно приходила к этому, независимо от эволюции не являющихся их предками групп прокариот. Скажем, происходила конвергенция на молекулярном уровне.
Симбиотическая гипотеза в последнее время находит большое коли-чество сторонников и часто называется уже не гипотезой, требующей дока-зательств, а теорией .
Следует отметить, что впервые идея о симбиотическом происхождении сложно устроенной клетки была высказана в России отечественным учёным-ботаником Андреем Сергеевичем Фаминцыным (1835-1918) в конце 60-х гг. XIX в. на основе изучения лишайников. В дальнейшем, в 1905-1909 гг., биолог Константин Сергеевич Мережковский (1855-1921) предложил тер-мин «симбиоз» и выдвинул концепцию о симбиотическом происхождении носителей пигментов в растительных клетках. Эта идея была развита в 1921-1924 гг. в симбиотическую концепцию происхождения клеток расте-ний и животных ботаником Борисом Михайловичем Козо-Полянским (1890-1957). На Западе эту концепцию развивали Лили Саган (1967), утверж-давшая, что эукариотическая клетка возникла путём симбиоза двух или более прокариотических клеток, Линна Маргулис (1983) и другие учёные.
Симбиотическая, или эндосимбиотическая, гипотеза исходит из того, что эукариоты представляют собой результат симбиоза между различными прокариотами . Допускается, что ядро развилось вследствие обособления ДНК из цитоплазмы путём впячивания плазматической мембраны и обвола-кивания ядерного вещества. Сходным образом могли возникнуть и эндоплазматическая сеть, и аппарат Гольджи .
Однако считается, что появление митохондрий и хлоропластов произо-шло иным путём. Полагают, что некоторые клетки, будучи гетеротрофами, за-хватывали других, более мелких бактерий , которых они по неизвестным при-чинам не переваривали. Захваченные мелкие клетки прокариот были способ-ны поглощать кислород. Это свойство было выгодным фактом для клетки-хозяина, поглотившей такую бактерию, так как давало 34 молекулы АТФ на одну молекулу глюкозы (против только двух молекул АТФ при брожении). По-глотителем могла быть и фототрофная клетка прокариот, питающаяся орга-ническими веществами, у которой в процессе реакций обмена веществ мог образовываться молекулярный кислород, ядовитый для неё, но потребляемый клеткой-симбионтом для своего дыхания. Из этих сохранившихся бактерий-симбионтов в ходе дальнейшей эволюции и могли произойти митохонд-рии. В пользу этой гипотезы свидетельствуют данные о современных симбио-тических отношениях между водорослями и инфузориями. На рис. 58 представлена схема возникновения в процессе эволюции сложной клетки эукари-от с помощью разных путей симбиоза. Материал с сайта
Одновременно с этими процессами или несколько позднее таким же путём мог произойти и эндоцитозный захват автотрофных прокариот, напри-мер сине-зелёных водорослей (цианобактерий), обладающих полным набо-ром пигментов фотосинтеза. Из этих симбионтов развились хлоропласты. Допускается также, что некоторые сложные по строению клетки, например простейших, произошли в результате симбиотических связей не только с про-кариотами, но и с другими эукариотами.
Симбиотическая гипотеза в настоящее время находит подтверждение рядом эволюционных реликтов в существующих и сейчас симбиотических связях между прокариотами и эукариотами. Такие связи наблюдаются сре-ди растительных и животных организмов. Например, свободноживущая амёба Pelomyxa palustris (пеломикса болотная) не имеет митохондрий, но содержит в себе симбиотические бактерии, которые обеспечивают ей ды-хание и АТФ.
На этой странице материал по темам:
Митохондрии - верные спутники эукариот. Согласно теории симбиогенеза, именно обретение митохондрий спровоцировало формирование ядерных организмов. Одним из доказательств этой теории было обнаружение митохондрий или подобных им органелл у всех, даже самых простых, эукариот. Но в мае 2016 года коллектив чешских ученых описал первый в истории ядерный организм, не содержащий даже косвенных признаков митохондрий. Может ли это открытие пошатнуть современные представления о ранней эволюции эукариот?
Митохондрии - двумембранные органеллы, снабжающие энергией клетки практически всех эукариот. Достоверно известно, что они родственны α-протеобактериям и стали частью эукариотических клеток около 1,5 млрд лет назад . О бактериальном происхождение митохондрий свидетельствует наличие двух мембран (внутренней собственной и внешней хозяйской), собственных кольцевой ДНК и трансляционной машины, а также способности независимо делиться. Некоторые даже полагают, что апоптоз - попытка митохондрии убить поглотившего ее эксплуататора.
Тем не менее в научном сообществе до сих пор нет единого мнения относительно роли этого симбиоза в развитии эукариот (рис. 1). Сторонники теории симбиогенеза утверждают, что слияние некой археи с предком митохондрий (бактерией) запустило цепочку событий, приведших к образованию эукариот современного типа. Приверженцы архезойной теории (гипотезы) , наоборот, считают, что «приручить» митохондрию мог только уже оформившийся ядерный организм - архезой .
Благодаря Карлу Вёзе и Джорджу Фоксу, в 70-х годах прошлого века сравнившим гены 16S рРНК множества живых существ, долго обманывавшие микробиологов своим визуальным сходством две формы доядерных организмов (прокариот) развели окончательно и бесповоротно, да еще и на высшем уровне: эубактерий лишили права на «истинность» (-эу), архебактерий - права носить гордое имя бактерий, зато в новой системе живых организмов им отвели по собственному домену (таксону высшего ранга): «Эволюция между молотом и наковальней, или как микробиология спасла эволюцию от поглощения молекулярной биологией » и «Карл Вёзе (1928–2012) » . Так в 1990 году человеку предложили осознать, что все живые существа филогенетически разбиваются на три домена: Бактерии, Археи и Эукариоты, - причем бактерии отличаются от архей даже больше, чем археи от эукариот, а недавно вообще чуть ли не усомнились в целесообразности разделения последних: «Нашли предков всех эукариот » . Однако предложение почти тридцатилетней давности до сих пор не нашло понимания у многих авторов отечественных учебников биологии. А что? Вдруг опять эти ученые всё поменяют, а им переписывать каждые -цать лет учебники что-ли? - Ред.
В 1928 году знаменитый биолог рубежа XIX-XX веков Эдмунд Вилсон высказался о гипотезе бактериального происхождения митохондрий так: «Подобные идеи чересчур фантастичны, чтобы их можно было обсуждать в приличном биологическом обществе» . Сегодня же подобное отношение сформировалось к архезойной теории, а ключевая роль митохондрий в ранней эволюции эукариот общепризнанна. Открытие первого истинно безмитохондриального простейшего заставляет еще раз задуматься о сильных и слабых сторонах каждой из теорий.
Теория симбиогенеза
Одно из преимуществ симбиогенетической теории по сравнению с архезойной в том, что она объясняет возникновение ядра и интронную структуру генома. У прокариот широко распространен горизонтальный перенос генов (ГПГ), за счет которого популяции могут быстро обмениваться частями генофонда . ГПГ способствует незащищенности генома - ведь поступающая ДНК в этом случае ничем не отделена от содержимого хозяйской клетки.
Вполне вероятные попытки клетки-хозяина разрушить пока еще не одомашненного симбионта приводили к высвобождению в цитоплазму симбиотической ДНК. Эта ДНК, будучи в непосредственной близости от хозяйского генома, могла легко в него встраиваться . Из-за ГПГ даже в эукариотах, утративших митохондрии, находят изначально митохондриальные гены.
Такое слияние геномов могло, во-первых, способствовать развитию взаимозависимости симбионта и хозяина. Во-вторых, обильный ГПГ мог переносить не только гены, обеспечившие переплетение метаболизма двух организмов, но и эгоистичные ретроэлементы . Вторжение интронов II группы, вырвавшихся из α-протеобактерий, привело к разрыхлению исходно очень плотного генома хозяина: до 80% хозяйской ДНК теперь были интронами , . В такой сложной ситуации клетка-хозяин развила несколько линий защиты своего генома от шквала интронов: возникли система внутренних мембран и ядро, убиквитиновая система деградации поврежденных белков, нонсенс-опосредованный распад РНК и прочие характерные особенности эукариот (рис. 3).
Рисунок 3. Формирование основных черт эукариот можно объяснить последовавшим за митохондриальным симбиозом вторжением интронов II группы в геном клетки-хозяина.
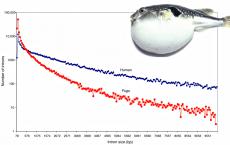
Еще одно мощное доказательство симбиогенеза - энергетические потребности эукариот. Хотя энергопотребление про- и эукариот в пересчете на грамм веса примерно одинаково, ядерные клетки гораздо крупнее безъядерных, из-за чего они потребляют примерно в 5000 раз больше энергии (2300 пВт/кл против 0,5 пВт/кл). При пересчете энергопотребления на средний ген одноклеточного организма оказывается, что эукариотический ген потребляет в 1000 раз больше энергии . Без митохондриальной энергетики было бы невозможно не только создать сложные, большие и активно передвигающиеся организмы, но даже обеспечить функционирование типичных для эукариот клеточных структур.
У гигантских бактерий масштабирование прокариотической энергетики за счет массовой полиплоидизации (как в случае Epulopiscium , дорастающей до 0,6 мм и содержащей 200 000 копий генома размером 3,8 млн п.н.) не приводит к повышению выхода энергии на ген, и клетка остается типично бактериальной . - Авт.
Еще один важный факт, подкрепляющий симбиогенетический сценарий - существование внутриклеточных симбионтов бактерий. Случаи эндосимбиоза у бактерий крайне редки в природе, но всё же они есть и демонстрируют, как мог зарождаться эукариотический домен жизни .
Архезойная теория
Архезои - предполагаемые безмитохондриальные, но ядерные предки современных эукариот. Согласно архезойному сценарию митохондрии были одомашнены только на поздних стадиях эволюции эукариот и не оказали на этот процесс значительного влияния.
Одно из основных положений симбиогенеза - гипотеза исходной простоты. О жизни во времена протерозоя известно крайне мало, поэтому о ее устройстве существует множество часто взаимоисключающих предположений. Если по первой гипотезе считается, что от прокариот с очень компактными геномами произошли более сложные эукариоты, то в архезойном сценарии изначально существовали клетки с запутанными и громоздкими геномами, от которых путем редукции произошли более простые прокариоты. Эукариоты же лишь сохранили первичную сложность.
Эволюция геномов и правда далеко не всегда движется от простого к сложному. И среди эукариот есть примеры, подтверждающие это.
Тем не менее редукция генома совсем не обязательно сопровождает его компактизацию. Доказательства этого можно найти как у простейших, так и у многоклеточных форм жизни.
Например, свободноживущая инфузория Paramecium tetraurelia содержит 30000 генов, на каждый из которых приходится в среднем 2 т.п.н. Такая компактность достигается за счет сокращения размера интронов до предельных 25 п.н. и уменьшения межгенных расстояний .
Даже у позвоночных могут быть необычно компактные геномы: геном рыбы фугу в восемь раз меньше человеческого в основном за счет низкого содержания повторов (рис. 4) .

Рисунок 4. Рыба фугу имеет необычно компактный для позвоночного геном частично благодаря коротким интронам. Вертикальная ось дана в логарифмическом масштабе.

Рисунок 5. Гипотеза изначальной сложности подразумевает, что прокариотические ветви жизни произошли от более сложных форм за счет редукции. Редукционный вектор развития мог быть задан первыми хищными архезоями, угнетающими прочие организмы.
Приведенные примеры показывают, что простота прокариотических геномов может возникать вторично. Если это так, то LUCA - последний общий предок всех современных организмов - мог обладать геномом эукариотического типа.
Гипотезу исходной сложности также подтверждают так называемые гены-сигнатуры («подписи») - эукариотические гены, не имеющие прокариотических гомологов. Вероятнее всего, эти гены содержались в LUCA, но были утеряны бактериями и археями.
К несчастью архезойной теории, список сигнатур значительно поредел с начала XXI века. Среди множества секвенированных с тех пор геномов были найдены их прокариотические гомологи. Таким образом, с каждым годом становится всё больше белков, чье присутствие у эукариот можно объяснить тем, что их гены были принесены архейным либо бактериальным предком в ходе симбиогенеза.
И в то же время открытие прокариотических гомологов белков клеточного движения (актинов, тубулинов и кинезинов) косвенно подтверждает возможность того, что архезои могли активно передвигаться и даже быть первыми хищниками на Земле, способными к фагоцитозу . Возникновение хищников в автотрофно-сапротрофном сообществе колыбели жизни должно было оказать колоссальное влияние на ход эволюции. В самых захватывающих сценариях одни жертвы архезоя приспосабливаются к быстрому делению и росту, а другие - к нишам, куда архезой не способен проникнуть. В итоге гипотетический архезой повел эволюцию своих современников по редуктивному пути с упором на гибкость метаболизма и скорость деления, в ходе чего сформировались известные нам прокариоты (рис. 5) .
Но несмотря на то, что архезойная теория имеет некоторые сильные стороны или как минимум наносит колкие удары в сторону симбиогенеза, ей не хватает главного - она не объясняет, как и почему сформировалось ядро .
Уникальная находка
В 1980-х существовало множество претендентов на звание современного архезоя, но в последующие годы у всех них нашли митохондриеподобные органеллы (митосомы и пероксисомы) и гены-маркеры митохондриального прошлого: гены сборки Fe-S -белков, митохондриальных транспортеров и шаперонов, синтетазы кардиолипина. К тому же некоторые белки, синтезируемые в цитоплазме, обладают последовательностями импорта в митохондрии, которые могут сохраняться и в отсутствие самих митохондрий.
С каждым новым «закрытием» потенциального архезоя безмитохондриальный сценарий становления эукариот оказывался всё менее вероятным. И вот в мае 2016 года наконец-то появился новый потенциальный архезой, не содержащий даже следов митохондрий.
Речь идет об анаэробной оксимонаде Monocercomonoides sp. PA203, живущей в кишечнике насекомых. Оксимонады лишены митохондрий и не содержат в ядерной ДНК гены митохондриального происхождения. Энергию они получают из гликолиза , идущего в цитоплазме.
Геном Monocercomonoides sp., расшифрованный коллективом чешских ученых, содержит 16629 генов, среди которых нет названных выше маркеров. Поиски митохондриальных гомологов и белков с импортными последовательностями тоже не дали удовлетворительных результатов (рис. 6).
Единственное, что удалось найти - два гена, продукты которых у близкого родственника Monocercomonoides sp. могут содержаться (а могут и не содержаться) в митохондриях, при этом они лишены импортных последовательностей.
Авторы открытия считают, что Monocercomonoides когда-то всё же содержали митохондрии, поскольку у близкородственных родов есть следы митохондрий. Еще остается возможность, что у этих простейших есть пока не обнаруженные митосомы, деградировавшие настолько, что в геноме не осталось каких-либо признаков их присутствия.
Так или иначе, Monocercomonoides sp. - пока уникальный случай истинно безмитохондриального протиста за всю историю биологии. И этот случай доказывает, что эукариоты могут жить не только без митохондрий, но и без их генетического наследства .
В какой последовательности предки эукариот разживались внутриклеточным скарбом и что стало счастливым билетом в эволюционное будущее, можно узнать из статьи «Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот » . - Ред.
Это открытие, конечно, не наносит сокрушительного удара по теории симбиогенеза, но однозначно заставляет задуматься, что есть необходимость и что есть излишество в эукариотах.
Литература
- Как появились митохондрии (рассказ, похожий на сказку) ;
- Кунин Е.В. Логика случая . М.: Центрполиграф, 2014. - 527 с.;
- Эволюция между молотом и наковальней, или Как микробиология спасла эволюцию от поглощения молекулярной биологией ;
- Карл Вёзе (1928–2012) ;
- Кондратенко Ю. (2015). «Нашли предков всех эукариот ». «Кот Шрёдингера» . 6 ;
- van der Giezen M. (2009).
Теория симбиогенеза (симбиотическая теория, эндосимбиотическая теория, или теория эндосимбиоза) объясняет механизм возникновения некоторых органоидов эукариотической клетки - митохондрий, пластид и гидрогеносом.
Суть концепция заключается в взаимовыгодном сожительстве органеллы с клеткой. Это позволяет предположить об эндосимбиозе, как о выгодном для обоих сторон симбиозе с образованием клеток эукариот (клетки, в которых присутствует ядро). Затем при помощи передачи генетической информации между бактериями осуществлялось их развитие и увеличение популяции. Согласно этой версии, все дальнейшие развитие жизни и жизненных форм обязано предшествующему предку современных видов.Теорию эндосимбиотического происхождения хлоропластов впервые предложил в 1883 году Андреас Шимпер, показавший их саморепликацию внутри клетки.
Ее возникновению предшествовал вывод А. С. Фаминцына и О. В. Баранецкого о двойственной природе лишайников - симбиотического комплекса гриба и водоросли (1867 год).
Четкие положения системы были составлены русским ботаником и зоологом К. С. Мережковским.
В 1905 году он предложил само название «симбиогенез», впервые детально сформулировал теорию и создал на ее основе новую систему органического мира. Фаминцын в 1907 году, опираясь на работы Шимпера, также пришел к выводу, что хлоропласты являются симбионтами, как и водоросли в составе лишайников.
В 1920-е годы теория была развита Б. М. Козо-Полянским, было высказано предположение, что симбионтами являются и митохондрии. Затем долгое время о симбиогенезе практически не упоминали в научной литературе. Второе рождение расширенная и конкретизированная теория получила уже в работах Линн Маргулис, начиная с 1960-х годов.
В результате изучения последовательности оснований в митохондриальной ДНК были получены весьма убедительные доводы в пользу того, что митохондрии - это потомки аэробных бактерий (прокариот), родственных риккетсиям, поселившихся некогда в предковой эукариотической клетке и «научившимися» жить в ней в качестве симбионтов. Теперь митохондрии есть почти во всех эукариотических клетках, размножаться вне клетки они уже не способны.
Существуют свидетельства того, что первоначально эндосимбиотические предки митохондрий не могли ни импортировать белки, ни экспортировать АТФ. Вероятно, первоначально они получали от клетки-хозяина пируват, выгода для хозяина состояла в обезвреживании аэробными симбионтами токсичного для нуклеоцитоплазмы кислорода.
Пластиды, подобно митохондриям, имеют свои собственные прокариотические ДНК и рибосомы. По-видимому, хлоропласты произошли от фотосинтезирующих бактерий, поселившихся в свое время в гетеротрофных клетках протистов и превратив их в автотрофные водоросли.
Доказательства
Митохондрии и пластиды:
Имеют две полностью замкнутые мембраны. При этом внешняя сходна с мембранами вакуолей, внутренняя - бактерий,
Размножаются бинарным делением (причем делятся иногда независимо от деления клетки), никогда не синтезируются de novo,
Генетический материал - кольцевая ДНК, не связанная с гистонами (По доле ГЦ ДНК митохондрий и пластид ближе к ДНК бактерий, чем к ядерной ДНК эукариот),
Имеют свой аппарат синтеза белка - рибосомы и другие рибосомы прокариотического типа - c константой седиментации 70S. По строению 16s рРНК близки к бактериальной.
Некоторые белки этих органелл похожи по своей первичной структуре на аналогичные белки бактерий и не похожи на соответствующие белки цитоплазмы.
Проблемы
ДНК митохондрий и пластид, в отличие от ДНК большинства прокариот, содержат интроны.
В собственной ДНК митохондрий и хлоропластов закодирована только часть их белков, а остальные закодированы в ДНК ядра клетки. В ходе эволюции происходило «перетекание» части генетического материала из генома митохондрий и хлоропластов в ядерный геном. Этим объясняется тот факт, что ни хлоропласты, ни митохондрии не могут более существовать (размножаться) независимо.
Не решен вопрос о происхождении ядерно-цитоплазматического компонента (ЯЦК), захватившего прото-митохондрии. Ни бактерии, ни археи не способны к фагоцитозу, питаясь исключительно осмотрофно. Молекулярно-биологические и биохимические исследования указывают на химерную архейно-бактериальную сущность ЯЦК. Как произошло слияние организмов из двух доменов, также не ясно.
Примеры эндосимбиозов
В наши дни существует ряд организмов, содержащих внутри своих клеток другие клетки в качестве эндосимбионтов. Они, однако, не являются сохранившимися до наших дней первичными эукариотами, у которых симбионты еще не интегрировались в единое целое и не потеряли своей индивидуальности. Тем не менее, они наглядно и убедительно показывают возможность симбиогенеза.
Mixotricha paradoxa - наиболее интересный с этой точки зрения организм. Для движения она использует более 250 000 бактерий Treponema spirochetes, прикрепленных к поверхности ее клетки. Митохондрии у этого организма вторично потеряны, однако внутри его клетки есть сферические аэробные бактерии, заменяющие эти органеллы.
Амебы рода Pelomyxa также не содержат митохондрий и образуют симбиоз с бактериями.
Инфузории рода Paramecium постоянно содержат внутри клеток водоросли, в частности, Paramecium bursaria образует эндосимбиоз с зелеными водорослями рода хлорелла (Chlorella).
Одноклеточная жгутиковая водоросль Cyanophora paradoxa содержит цианеллы - органоиды, напоминающие типичные хлоропласты красных водорослей, однако отличающиеся от них наличием тонкой клеточной стенки, содержащей пептидогликан. Размер генома цианелл такой же, как у типичных хлоропластов, и во много раз меньше, чем у цианобактерий.
Гипотезы эндосимбиотического происхождения других органелл
Эндосимбиоз - наиболее широко признанная версия происхождения митохондрий и пластид. Но попытки объяснить подобным образом происхождение других органелл и структур клетки не находят достаточных доказательств и наталкиваются на обоснованную критику.
Пероксисомы
Кристиан де Дюв обнаружил пероксисомы в 1965 году. Ему же принадлежит предположение, что пероксисомы были первыми эндосимбионтами эукариотической клетки, позволившими ей выживать при нарастающем количестве свободного молекулярного кислорода в земной атмосфере. Пероксисомы, однако, в отличие от митохондрий и пластид, не имеют ни генетического материала, ни аппарата для синтеза белка. Было показано, что эти органеллы формируются в клетке de novo в ЭПР и нет никаких оснований считать их эндосимбионтами.
Расцвет эукариот на Земле начался около 1 млрд лет назад, хотя первые из них появились намного раньше (возможно 2,5 млрд лет назад). Происхождение эукариот могло быть связано с вынужденной эволюцией прокариотических организмов в атмосфере, которая стала содержать кислород.
Симбиогенез - основная гипотеза происхождения эукариот
Существует несколько гипотез о путях возникновения эукариотических клеток. Наиболее популярная - симбиотическая гипотеза (симбиогенез) . Согласно ей, эукариоты произошли в результате объединения в одной клетке разных прокариот, которые сначала вступили в симбиоз, а затем, все более специализируясь, стали органоидами единого организма-клетки. Как минимум симбиотическое происхождение имеют митохондрии и хлоропласты (пластиды вообще). Произошли они от бактериальных симбионтов.
Клеткой-хозяином мог быть относительно крупный анаэробный гетеротрофный прокариот, похожий на амебу. В отличие от других, он мог приобрести способность питаться путем фаго- и пиноцитоза, что позволяло ему захватывать других прокариот. Они не все переваривались, а снабжали хозяина продуктами своей жизнедеятельности). В свою очередь, получали от него питательные вещества.
Митохондрии произошли от аэробных бактерий и позволили клетке-хозяину перейти к аэробному дыханию, которое не только намного эффективней, но и облегчает существование в атмосфере, содержащей достаточно большое количество кислорода. В такой среде аэробные организмы получают преимущество над анаэробными.
Позже в некоторых клетках поселились похожие на ныне живущих синезеленых водорослей (цианобактерий) древние прокариоты. Они стали хлоропластами, дав начало эволюционной ветви растений.
Кроме митохондрий и пластид симбиотическое происхождение могут иметь жгутики эукариот. В них превратились симбионты-бактерии наподобие современных спирохет, имеющих жгутик. Считается, что в последствии из базальных тел жгутиков произошли центриоли, столь важные структуры для механизма клеточного деления эукариот.
Эндоплазматическая сеть, комплекс Гольджи, пузырьки и вакуоли могли произойти от наружной мембраны ядерной оболочки. С другой точки зрения, некоторые из перечисленных органелл могли возникнуть путем упрощения митохондрий или пластид.
Во многом неясным остается вопрос происхождения ядра. Могло ли оно также образоваться из прокариота-симбионта? Количество ДНК в ядре современных эукариот во много раз превышает его количество в митохондриях и хлоропластах. Возможно часть генетической информации последних со временем переместилась в ядро. Также в процессе эволюции происходило дальнейшее увеличение размера ядерного генома.
Кроме того в симбиотической гипотезе происхождения эукариот не все так однозначно с клеткой-хозяином. Им мог и не быть один вид прокариот. Используя методы сравнения геномов, ученые делают вывод, что клетка-хозяин близок к археям, при этом сочетает в себе признаки архей и ряда неродственных групп бактерий. Отсюда можно сделать вывод, что появление эукариот происходило в сложном сообществе прокариот. При этом процесс скорее всего начался с метаногенной археи, вступавшей в симбиоз с другими прокариотами, что было вызвано необходимостью обитания в кислородной среде. Появление фагоцитоза способствовало притоку чужих генов, а ядро образовалось для защиты генетического материала.
Молекулярный анализ показал, что различные белки эукариот происходят от разных групп прокариот.
Доказательства симбиогенеза
В пользу симбиотического происхождения эукариот говорит то, что митохондрии и хлоропласты имеют собственную ДНК, причем кольцевую и не связанную с белками (также обстоит дело у прокариот). Однако в генах митохондрий и пластид есть интроны, чего нет у прокариот.
Пластиды и митохондрии не воспроизводятся клеткой с нуля. Они образуются из ранее существующих таких же органелл путем их деления и последующего роста.
В настоящее время существуют амебы, у которых нет митохондрий, а вместо них есть бактерии симбионты. Также есть простейшие, сожительствующие с одноклеточными водорослями, выполняющими в клетке-хозяине роль хлоропластов.

Инвагинационная гипотеза происхождения эукариот
Кроме симбиогенеза существуют и другие взгляды на происхождение эукариот. Например, инвагинационная гипотеза . Согласно ей, предком эукариотической клетки был не анаэробный, а аэробный прокариот. К такой клетке могли прикрепляться другие прокариоты. Потом их геномы объединялись.
Ядро, митохондрии и пластиды возникли путем впячивания и отшнуровывания участков клеточной мембраны. В эти структуры попадала чужеродная ДНК.
Усложнение генома происходило в процессе дальнейшей эволюции.
Инвагинационная гипотеза происхождения эукариот хорошо объясняет наличие двойной мембраны у органелл. Однако она не объясняет, почему система биосинтеза белка в хлоропластах и митохондриях сходна с прокариотической, в то время как таковая в ядерно-цитоплазматическом комплексе имеет ключевые отличия.
Причины эволюции эукариот
Все разнообразие жизни на Земле (от простейших до покрытосеменных и млекопитающих) дали клетки эукариотического, а не прокариотического типа. Возникает вопрос, почему? Очевидно, ряд особенностей, возникших у эукариот, существенно повысили их эволюционные возможности.
Во-первых, у эукариот есть ядерный геном, который во много раз превосходит количество ДНК у прокариот. При этом эукариотические клетки диплоидны, кроме этого в каждом гаплоидном наборе определенные гены многократно повторяются. Все это обеспечивает, с одной стороны, большие масштабы для мутационной изменчивости, а с другой - уменьшает угрозу резкого снижения жизнеспособности в результате вредной мутации. Таким образом, эукариоты, в отличие от прокариот, обладают резервом наследственной изменчивости.
Эукариотические клетки имеют более сложный механизм регуляции жизнедеятельности, у них существенно больше различных регуляторных генов. Кроме того, молекулы ДНК образовали комплексы с белками, что позволило наследственному материалу упаковываться и распаковываться. Все вместе это дало возможность считывать информацию частями, в разных сочетаниях и количестве, в разное время. (Если в клетках прокариот транскрибируется почти вся информация генома, то в эукариотических клетках обычно менее половины.) Благодаря этому эукариоты могли специализироваться, лучше приспосабливаться.
У эукариот появились митоз, а затем и мейоз. Митоз позволяет воспроизводить генетически сходные клетки, а мейоз сильно увеличивает комбинативную изменчивость, что ускоряет эволюцию.
Большую роль в процветании эукариот сыграло приобретенное их предком аэробное дыхание (хотя оно есть и у многих прокариот).
На заре своей эволюции эукариоты обзавелись эластичной оболочкой, обеспечивавшей возможность фагоцитоза, и жгутиками, позволившими им двигаться. Это дало возможность эффективней питаться.